|
Humberside Geologist No.12
Copyright Hull Geological Society 1999.
Rudist bivalves - a bizarre group of fossils from the Cretaceous of Jamaica
Simon F. Mitchell
Department of Geography and Geology, University of the West Indies, Mona,
Kingston., Jamaica
Introduction
The end-Cretaceous mass extinction saw the demise of many fossil groups including the dinosaurs, ammonites and belemnites. One of the most bizarre groups (both in growth form and size) to disappear near the end of the Cretaceous was the rudist bivalves, which had dominated tropical carbonate platforms since the late Jurassic. Although abundant in Cretaceous tropical deposits, the group is poorly represented in most textbooks on palaeontology.
Rudists are anomalous bivalves which developed massive sessile shells, often with the two valves showing strong asymmetry (Dechaseaux et al., 1969), and were adapted to filter feeding. Many forms show the strong development of the attached valve (AV) and the subsequent reduction of the free valve (FV). In general form, rudists resemble corals and it has been suggested that rudists displaced corals from reefs during the Cretaceous (Kauffman and Sohl 1974; Johnson and Kauffman, 1996).
In Jamaica rudists are abundant and were one of the major carbonate sediment producers in rocks of Cretaceous age. They are therefore important in studying ancient tropical environments. Their peculiar, and often bizarre form also make them an interesting curiosity.
Rudists in Jamaica
Rudists were discovered in Jamaica by the young Englishman Lucus Barrett who was the director of the first geological survey of Jamaica. Barrett found a large species in the northern Blue Mountains which he recognised as a species of a hippurite rudist (Barrett, 1860), and thus demonstrated that rocks previously thought to be of Palaeozoic age (De la Beche, 1827) were of Cretaceous age. The specimens were returned to London and named Barrettia in honour of the young geologist by Samuel Thomas Woodward (1862). Subsequent rudists were collected by Nicholas in 18951896 and described by Whitfield (1897a, b). Another Englishman who became interested in Caribbean geology was C. T. Trechmann who collected extensively from the rocks of Jamaica and published a succession of works in which he described rudists as well as other fossils (Trechmann, 1922, 1924). With the establishment of a permanent Geological Survey Department in Jamaica in the 1950s, Lawrence Chubb moved to Jamaica. He became interested in the rudists and produced many papers revising the fauna (Chubb, 1955, 1956a, b, c); this culminated in the appearance of his monograph 'Rudists of Jamaica', which was published in 1971 shortly before his death.
Classification of rudist morphotypes
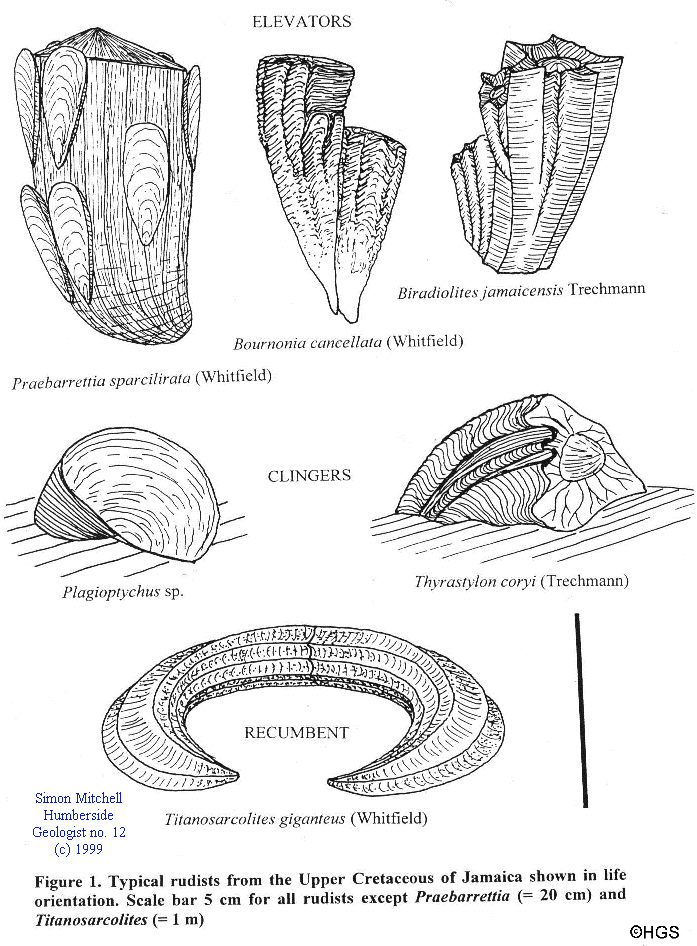
Skelton and Gilli (1991) have suggested a morphotype classification for rudist shells based on their life orientation (this is not an evolutionary classification since the same growth plan developed separately in many unrelated groups of rudist). They recognised three morphotypes: elevators, clingers and recumbents.
Elevators (Fig. 1) grew with a vertical to subvertical growth pattern, they thus maintained their commissure almost parallel to the sediment-water interface. Their attached valves are generally conical to cylindrical with a small basal attachment area, while their free valves may be coiled or reduced to a small cap. Elevators grew either singularly, in close associations or in dense clusters with mutual cementation. Although some reconstructions show elevators largely buried within the sediment (e.g., Ross & Skelton, 1991), the distribution of epibiont (encrusting) and boring organisms on the shells of Caribbean elevators (Kauffman and Sohl, 1974) suggests that most elevators were only slightly, if at all, buried within the sediment. The same can be determined from the preservation of small clusters of in situ elevators within sediments in Jamaica. Elevators are the most diverse rudists that occur within the Cretaceous deposits of Jamaica ranging from small forms (such as Bournonia) a few centimetres long to giant barrel shaped forms (such as Barrettia) which may be up to 1 m long and 50 cm or more in diameter.
The clingers (Fig. 1) were attached by a complete side of the attached valve. They have a commissure which is inclined at a moderate angle to the sediment-water interface. Clingers largely occur as single specimens attached to hard substrates such as corals, hardgrounds and other rudists. The most distinctive clinger in Jamaica is Plagioptychus.
The final morphological group is the recumbents (Fig. 1), which lie with the long axis of both valves parallel to the sediment-water interface. They commonly grew with horseshoe-like forms and had their commissure at 90' to the sedimentwater interface. The most distinctive species Titanosarcolites giganteus (Whitfield) with a length up to 2 m for complete specimens is amongst the largest of known molluscs!
The life of Jamaican rudists
Rudists in Jamaica occur in limestones interbedded with thick successions of volcaniclastic sediments and lavas. We can envisage episodes of prolonged volcanic activity with extensive andesitic volcanism associated with an ancient volcanic island arc, alternating with volcanically quiet interludes during which nudists colonised the shallow water around the dormant volcanoes. The carbonates also contain a wealth of corals, however the rudists and corals are largely exclusive, that is beds which contain abundant nudists contain few corals and vice versa. In fact, the beds with corals contain a fairly normal marine 'reef-type' fauna (e.g., algae, bryozoans, serpulids, gastropods, echinoids, starfish, crabs and brachiopods), while the beds with rudists contain a restricted fauna with only low diversity larger foraminiferal assemblages, scattered gastropods and one species of echinoid. Furthermore, the rudist limestones are also characterised by the presence of abundant mud-grade carbonate suggesting quiet water conditions, although the general lack of rudists in life position indicates either toppling after death or periodic reworking by storms. The difference in the faunas between the coral-rich and rudist-rich beds suggests that the corals grew, often as low-level patch reefs, in open marine waters, while the rudists inhabited more restricted quiet-water environments lacking extensive wave energy other than during storms.
Extinction
The end-Cretaceous mass extinction affected many groups of organisms, however, the rudists appear to have succumbed prior to the main extinction event. Since they inhabited shallow water carbonate environments, they were undoubtedly highly susceptible to sea level change, both rises (drowning platforms) and falls (exposing platforms). Rudists were, therefore, highly susceptible to sea-level change. Throughout the Caribbean, the limestones which contain rudists are succeeded by either volcanic deposits or deep-water turbidite sequences. This suggests that the rudist extinction in the Caribbean may have been caused by the break up and rapid subsidence of the various portions of the carbonate platform, rather than by the variously debated mechanisms that caused the demise of the Dinosaurs.
References
Barrett, L. 1860. On some Cretaceous rocks in the south-eastern portion of Jamaica.
Quarterly Journal of the Geological Society, 16, 324- 326
De la Beche, H.T. 1927. Remarks on the geology of Jamaica. Trans. Geol. Soc. London 2 nd. Ser., 2, no. 13:143-194.
Chubb, L. J. 1955. A revision of Whitfield's type specimens of the rudist mollusks from the
Cretaceous of Jamaica, B. W. 1. American Museum Novitates, No. 1713, 1-15.
Chubb, L. J. 1956a. Two rare rudists from Jamaica, B. W. 1. Palaeontographica Americana,
4, 1-30.
Chubb, L. J. 1956b. Thyrastylon, a new rudist genus from the Upper Cretaceous of Guatemala, the Antilles and Persia, with a discussion on the function of rudist oscules and pillars. Palaeontographica Americana, 4,31-48.
Chubb, L. J. 1956c. Rudist assemblages of the Antillean Upper Cretaceous. Bulletins of American Paleontology, 37, 5-3 1.
Chubb, L. J. 197 1. Rudists of Jamaica. Palaeontographica Americana, VII, No. 45, 161-25 7.
Dechaseaux, C., Coogan, A. H., Cox, L. R. and Perkins, B. F. 1969. In Treatise on invertebrate paleontology (Moore, R. C., ed.) part N, Mollusca 6 (Bivalvia) vol. 2, N776-N817, Lawrence, Kansas, Geological Society of America and University of Kansas.
Johnson, C. C. and Kauffman, E. K. 1996. Maastrichtian extinction patterns of Caribbean Province rudists. Chapter 9 in MacLeod, N. and Keller, G. Cretaceous-Tertiary mass extinctions: biotic and environmental changes. Norton and Company, New York-London, 23 1-272.
Kauffman, E. G. and Sohl, N. F. 1974. Structure and evolution of Antillean Cretaceous rudist frameworks. Verhandl. Naturf. Ges. Basel, 84,399-467.
Ross, D. J. & Skelton, P. W. 1993. Rudist formations of the Cretaceous: a palaeoecological, sedimenological and stratigraphical review.
In Sedimentology Review
1(V. P. Wright, Ed.), 73-91, Blackwells.
Skelton, P. W. and Gilli, E. 1991. Palaeoecological classification of rudist morphotypes.
Proceedings of the 1st International Conference on Rudists (Beograd, 1988), Serbian Geological Society, Special Publication
2, 265-287.
Trechmann, C. T. 1922. The Barrettia beds of Jamaica. Geological Magazine, 59, 501-514.
Trechmann, C. T. 1924. The Cretaceous limestones of Jamaica and their Mollusca.
Geological Magazine,
61, 385-410.
Whitfield, R. P. 1897a. Descriptions of species of Rudistae from the Cretaceous rocks of Jamaica, W. I., collected and presented by Mr. F. C. Nicholas. Bulletins of American Museum of Natural History, 9, 185-196. Whitfield, R. P. 1897b. Observations on the genus Barrettia Woodward, with descriptions of two new species. Bulletins of American Museum of Natural History, 9, 233-246. Woodward, S. P. 1862. Some account of Barrettia, a new and remarkable fossil shell from the Hippurite limestone of Jamaica. The Geologist 5, 372-377.
Copyright Hull Geological Society, (c) 1999
copyright Hull Geological Society 2020